序論
前々回(その2-1)では、赤色矮星セファエルとその惑星系の天体力学を設計した。潮汐加熱による磁場の維持、衛星系による離心率のポンプ、準同期自転——これらすべてが、薄暗く音声言語より光語が有利な舞台を作り出すための骨組みだった。
前回(その2-2)では、その舞台に「呼吸できる空気」を用意した。しかし結果は皮肉なものだった。温室効果を確保するため、二酸化炭素濃度は 5.0〜5.5% に達した。地球基準では重度の危険域だ。人間なら短時間で激しい息切れ・頭痛・意識混濁に至り、さらに高濃度(8–10%)では急速に意識を失う。産業安全衛生の現場では** 0.5% が長時間の上限目安、潜水艦でも運用上限は 0.8% 程度**である。
CO₂を減らせばいいのか? それは不可能だ。メタンは可燃性の壁(5%の爆発下限)があり、300 ppm以下に抑えざるを得ない。水蒸気は飽和蒸気圧の壁(熱力学の法則)により、柱平均0.9%が上限だ。残りの+43〜49°Cの温室効果を担えるのは、CO₂しかない。5%を割れば、惑星は凍りつく。
つまり、「温度的には住める」が「呼吸はできない」、そんな袋小路に追い込まれたように見える。
しかし、ここで立ち止まって考えてみよう。なぜ地球の生物はCO₂ 5%で死ぬのか? それは私たちが0.04%(400 ppm)という極低濃度環境に適応しているからだ。地球の大気組成は、約27億年前の大酸化イベント(シアノバクテリアによる酸素生産)以降、酸素優勢・CO₂劣勢という特殊な状態にある。しかしこれは宇宙的には例外だ。
実際、地球の初期生命(約 40 億年前)は、高 CO₂・低 O₂ の環境下で海から立ち上がった。大気の CO₂ 比率については推定に幅があり、時期と手法で 0.1%〜数%、あるいはそれ以上と議論が分かれるが、少なくとも現在より桁違いに高濃度だったことは確かだ。
しかし、問題は「どうやって適応するか」だ。そして、その適応戦略こそが、音声言語を使えなくする鍵になる。今回では、ルブラリスでの生物進化の歴史を、原始生物から始めて現代の知的生命に至るまで追う。特に焦点を当てるのが「呼吸器官の進化」だ。
地球の歴史を振り返ってみよう。最初の生命は海で生まれた。やがて酸素が増え、オゾン層ができ、陸上進出が可能になった。魚は鰓(えら)で水中の酸素を取り込んでいたが、一部は「浮き袋」を肺に転用し、空気呼吸を始めた。両生類、爬虫類、哺乳類——陸上動物はすべて、この肺を持つ。
しかし肺だけでは、まだ言語は生まれない。決定的だったのは「咽頭の獲得」だ。咽頭(いんとう)とは、口と食道・気管の分岐点にある空間だ。ここに声帯があり、空気の流れを調整することで複雑な音声を作り出せる。魚には咽頭がない。両生類や爬虫類にも、哺乳類ほど発達した咽頭はない。鳥類は独自の鳴管(めいかん)を持つが、構造が全く異なる。
つまり、言語の前提は二つだ:
- 肺による呼吸:空気を大量に出し入れできる器官
- 咽頭の獲得:その空気流を音声に変換する器官
地球では、この二つが揃うのに約4億年かかった。魚類の肺の原型が登場したのが約4.2億年前(デボン紀)、哺乳類の咽頭が発達したのが約2億年前(三畳紀後期)だ。
では、ルブラリスではどうか? CO₂ 5%という環境は、地球とは全く異なる進化の道筋を強いる。肺は獲得できるのか? 咽頭は形成されるのか? そして、もし形成されないとしたら——それが光語への鍵となる。
今回からルブラリスの呼吸の話がメインになる。本来はその2-3の一部として簡単に流し、発光器官に移るはずだったが、地上呼吸に至るまでのメカニズムがCO₂ 5%という環境のため大きく屈折することになった。そのため、新しく「その3」として章立てすることになった。申し訳ないが、光語、そしてそれをあやつる宇宙人の輪郭を捉えるためにもう少しお付き合いしてもらいたい。
鰓から始まる呼吸生活
植物が光合成で有機物を作り、酸素を放出する。その有機物と酸素を使って生きる生物。それが動物だ。
しかしルブラリスの動物は、地球の動物とは根本的に異なる制約を背負っていた。それが高CO₂大気だ。現在4%、初期には10%以上——これは地球の動物にとっては即死レベルだ。
この環境で呼吸するには、地球とは全く異なる呼吸器官が必要になる。その進化の道筋を、地球の例と比較しながら追っていこう。
地球の鰓
まず地球の魚類の鰓(えら)を理解しよう。鰓は単なる「酸素取り込み装置」ではない。いくつかの重要な機能を同時に果たしている。
酸素の取り込み
これが最も有名な機能だ。海水中に溶けている酸素(O₂)を血液に取り込む。
仕組み:
【鰓での酸素交換】
海水(溶存O₂:およそ5〜8 mg/L※温度・塩分で変動)
↓
鰓弁(極薄膜;毛細血管密集)
↓
血液(ヘモグロビンがO₂を結合)
鰓弁は数〜数十 µmの薄さで巨大な表面積を確保している。体重1 kgあたりの鰓表面積はおよそ1,000〜5,000 cm²/kg(種と活動性で幅)に達し、この広い界面が高効率のガス交換を支える。
鍵は対向流交換だ。血液と海水を逆方向に流すことで、鰓列のどの位置でも血液側の酸素分圧が海水側より低く保たれ、理論上80〜90%に迫る取り込み効率を実現する。
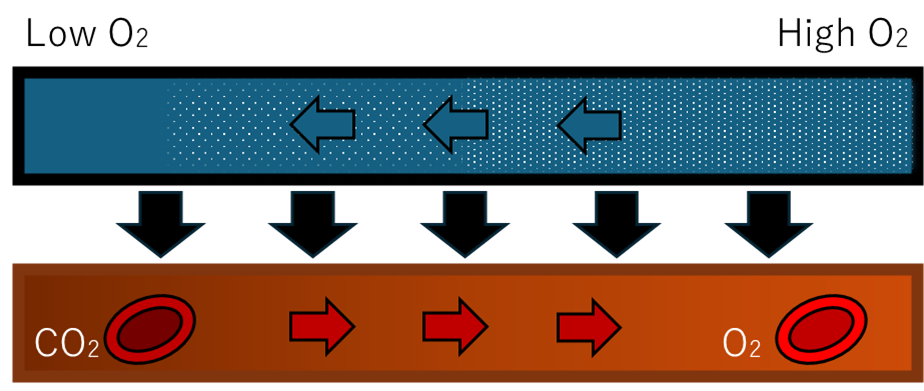
二酸化炭素の排出
呼吸(代謝)で生じたCO₂を、血液から海水へ排出する。同時に血液pHの維持(酸塩基調節)にも直結するため、生理的には「副次」と言い切れない重要任務だ。
仕組み:
地球の表層海は弱アルカリ(pH 8.1前後)、DIC 2〜3 mM。血液のpCO₂(約40 mmHg)は海水側の実効pCO₂より高いため、分圧勾配に従ってCO₂が自然に放散される。
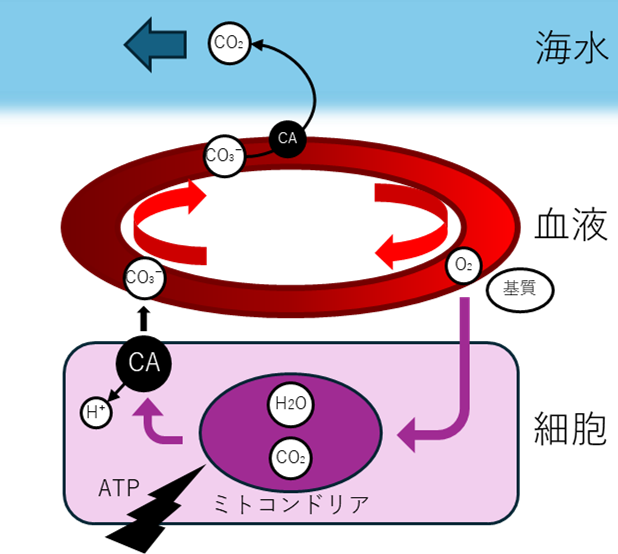
ただしCO₂の排出には、少し複雑な仕組みがある。血液中のCO₂の大部分は、実はCO₂分子のままではなく、炭酸水素イオン(HCO₃⁻)として運ばれている。CO₂が赤血球に入ると、CA(炭酸脱水酵素)というものがCO₂と水を結び付け、HCO₃⁻にして血漿に流す。
なぜこんな面倒なことをするのか? CO₂分子のままより、HCO₃⁻イオンの形の方が血液中に大量に溶かせるからだ。水に溶けるCO₂は限られているが、HCO₃⁻ならもっと多く運べる。
ここで重要なのは、最終的にCO₂分子として排出されるということだ。地球の海では、血液のpCO₂(40 mmHg)> 海水のpCO₂(0.3 mmHg)という圧倒的な濃度差がある。だから、あとはCAでもう一度CO₂に戻しさえすれば、あとは勝手に海水へ拡散していく。エネルギーを使って「汲み出す」必要はない。
ただし、HCO₃⁻を効率よく排出するため、鰓の細胞(塩類細胞。役割は後述)にはCl⁻/HCO₃⁻交換体(SLC26)というポンプも備わっている。名前の通り、細胞の内外でCl-とHCO₃-を交換する膜貫通タンパク質だ。
これは主に血液のpH調節に使われる。運動時など、大量のCO₂が発生すると血液が酸性に傾くため、HCO₃⁻を素早く排出してpHを保つ必要がある。しかし地球の魚にとって、これは「補助的」な機能だ。CO₂排出の主役は、あくまでCO₂分子の単純拡散である。
アンモニア(窒素老廃物)の排出
タンパク質代謝で生じる窒素は、水生脊椎動物では主にアンモニア(NH₃/NH₄⁺)として処理される。極めて毒性が高いが、水中という“大容量の受け皿”があるため、希釈して継続的に捨てられる。
鰓でのアンモニア排出の実像:
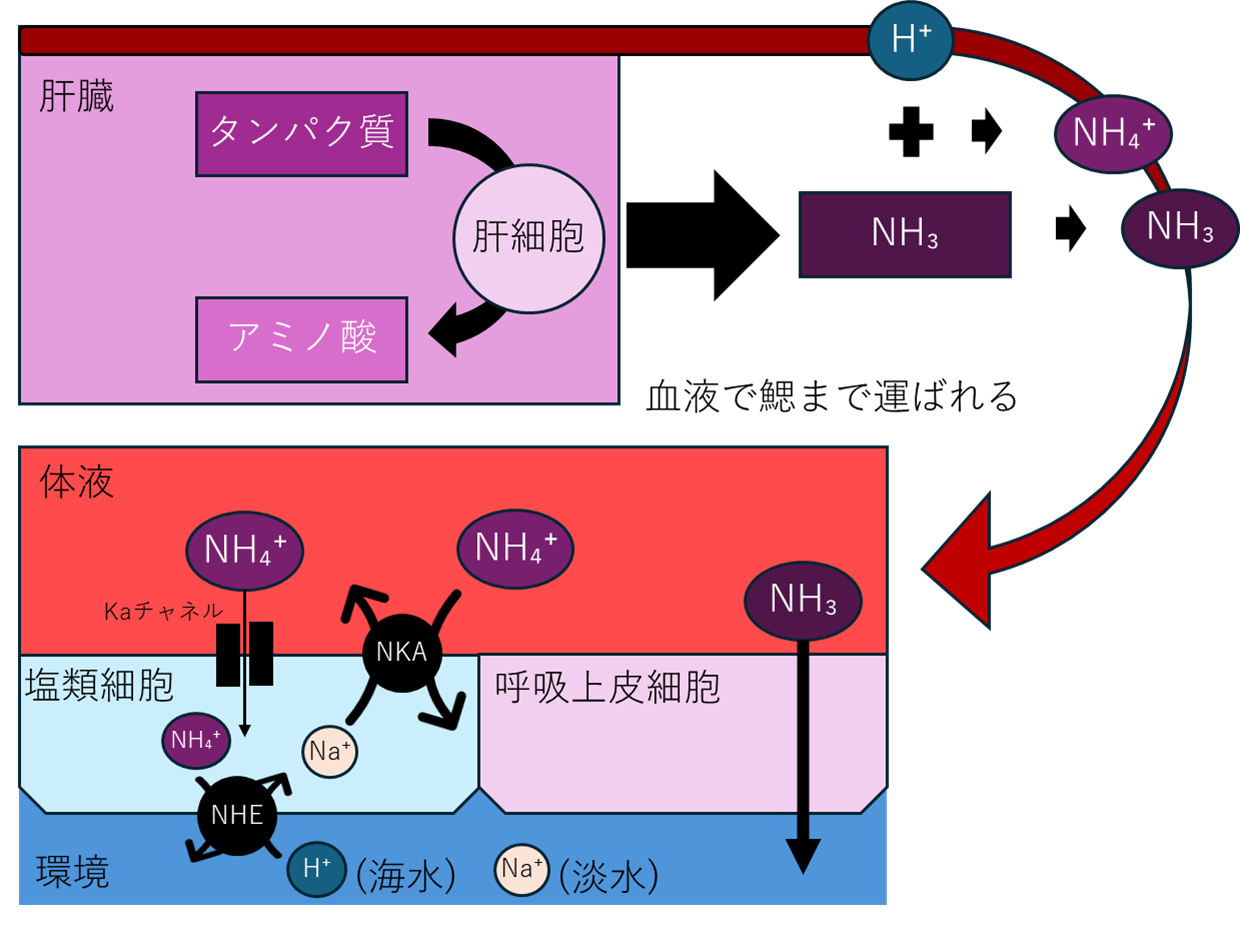
消化して吸収したタンパク質は、肝臓での代謝でアミノ酸ができると同時に、副産物でアンモニア(NH₃)が生成。血中に溶け込んで、アンモニウムイオン(NH₄⁺)になるものと分子のままのものの二つで存在し、これらの量はpH依存で平衡する。
NH₃は鰓上皮の呼吸上皮細胞、そして塩類細胞から直に排出される。NH₃は小さくて、極性が弱くて、脂質でできた細胞膜に溶け込みやすい。そのため、細胞膜を自由に通過できる。血液中のNH₃濃度は約0.01〜0.1 mM、海水中はほぼゼロ。この濃度差があるため、NH₃は勝手に海水へ拡散していく。エネルギー不要だ。
より専門的に言うならば、脂質二重層の単純拡散に加え、呼吸上皮細胞にあるRhesus(Rh)タンパク質と呼ばれるチャネルが選択的なNH₃透過を担うと考えられている。Rhチャネルは選択的にNH₃を通すトンネルのような構造で、拡散を3〜10倍速くする。しかし本質的には、これは拡散の効率化であって、能動輸送ではない。
一方、NH₄⁺(アンモニウムイオン)は、細胞膜を通過できない。NH₄⁺はプラスの電荷を持つイオンで、脂質の膜を通り抜けられないからだ。しかし血液中のアンモニアの99%はNH₄⁺の形なので、これも排出する必要がある。よって、NH₄⁺は、魚のあるメカニズムに相乗りして排出される。浸透圧調節だ。
体液と比べ、海水の場合は塩分濃度が高く、淡水の場合は塩分濃度が低い。そのため、鰓が水と直接触れると、放置すれば海水中では体内から水が奪われ、淡水中では過剰な水が流入し、いずれも致死的な浸透圧変化を引き起こす。よって、魚にはNHE(H⁺-ATPase)、NKA(Na+/K+-ATPase)という二つのポンプを備えた細胞で体の塩分濃度を一定に保つ。これを塩類細胞という。
塩類細胞は環境と体液を挟むバッファとして、NHEが環境向き、NKAが体液向きに働く。NHEは主にH⁺とNa⁺を交換する。一方でNKAは、ATPを使ってNa⁺とK⁺を交換する。これにより、塩類細胞は環境と体液の間でNa⁺・K⁺・H⁺の流れを制御し、内部のイオン濃度を動的に安定化させている。NH₄⁺はK⁺と性質が似ているため、このNa⁺/K⁺輸送経路を“借用”して体外へ排出される。
海水環境では、外界のNa⁺濃度が体液よりはるかに高く、浸透圧的にはNa⁺が体内に侵入しようとする。そのため、魚はNKAを高活性化させてNa⁺を能動的に排出し、同時に水分喪失を補うための飲水行動をとる。
このとき、NKAのK⁺輸送経路にはNH₄⁺が代替的に利用され、結果としてNa⁺排出と同時にアンモニア(NH₃/NH₄⁺)の排泄も進行する。
一方、淡水環境では、外界のNa⁺濃度が低く、逆に体内の塩分が流出しやすい。このためNHEが主に機能し、H⁺の放出と引き換えにNa⁺を取り込み、同時にNH₄⁺を体外へ排出する。環境のpHが中性〜弱酸性である場合、放出されたNH₄⁺は直ちにNH₃へと平衡変換し、拡散によって外界へ散逸する。
要するに、CO₂排出とNH₃排出は、H⁺の分泌・境界層pH制御という共通の基盤の上に、それぞれCA(CO₂系)とRhチャネル(NH₃系)が重なって動いている。鰓はガス交換・酸塩基調節・窒素排出・浸透圧調整を一体でこなす、高度に統合された界面臓器なのだ。
ルブラリスの鰓
ルブラリスの海洋環境は、地球とは全く異なる。ルブラリスの海は、高DIC・高CO₂分圧という極端な環境だ。それ故に、この環境で動物が進化すると、鰓の機能が逆転する。
ルブラリスの魚類相当が出現したのは大酸化イベントによってCO₂がある程度落ち着き、海洋化学が比較的安定した時期だ。この時代、大気CO₂は既に5〜6%程度まで低下し、表層海のpCO₂は140〜170 mmHg、pHは8.2〜8.5の範囲に収まっていた。極端なソーダ海洋期(CO₂ 10〜20%、pH 9.0〜9.5)は既に終わっており、魚類相当生物はこの「新常態」に最適化された形で進化を開始した。
しかし、「落ち着いた」とはいえ、地球の海(pCO₂ 0.3〜0.4 mmHg)と比較すれば依然として350〜425倍もの高CO₂環境だ。血液のCO₂分圧は代謝で生じた分を含めて約50〜100 mmHgと見積もると、この海では血液よりも海中の方が二酸化炭素が高い。
【地球とルブラリスの比較】
地球:
血液pCO₂(40 mmHg)>> 海水pCO₂(0.3 mmHg)
→ CO₂は勝手に海水へ拡散(エネルギー不要)
ルブラリス:
血液pCO₂(50〜100 mmHg)< 海水pCO₂(140〜170 mmHg)
→ CO₂は海水から血液へ逆流しようとする!
つまり、CO₂が勝手に排出されないどころか、海水から血液へ逆流する危険さえあるということだ。なぜか?拡散は常に濃度の高い方から低い方へ進むという物理法則があるからだ。
よって、ルブラリスではCA(炭酸脱水酵素)を地球の魚よりずっと多い濃度で鰓上皮に存在させなくてはならない。なぜならば、血液から入ってくるCO₂を、海水から侵入してくるCO₂と混ざる前に素早くHCO₃⁻に変換する必要があるからだ。
CAが不足すると、濃度勾配に従って鰓上皮細胞がCO₂の「通り道」になってしまい、海水の高濃度CO₂が血液に流れ込む危険がある。これを防ぐには、細胞内に入ったCO₂を即座にHCO₃⁻に変換して「捕獲」する必要がある。
HCO₃⁻はイオンなので、脂質でできた細胞膜を通過できない。つまり、一度HCO₃⁻に変換してしまえば、細胞内に閉じ込められる。そして、能動輸送ポンプで制御的に海水へ排出できる。
ルブラリスの鰓上皮細胞は、このCAを地球の魚の5〜8倍も詰め込むことになる。細胞質の5〜10%がCAで占められているとさえ推定される。これは異常な高濃度だ。
しかし、それ以上に問題がある。それはHCO₃⁻は陰イオンであることだ。
NH₃は分子なので、細胞膜を勝手に通り抜けられた。NH₄⁺は陽イオンだが、幸いK⁺と性質が似ていたため、既存のNKA(Na⁺/K⁺-ATPase)ポンプに相乗りできた。どちらも「専用の排出システム」を作る必要がなかったのだ。しかし、HCO₃⁻を排出するには、専用の輸送システムを作らなければならない。
さらに厄介なのは電荷のバランスだ。生物の細胞は、細胞膜の内外で一定の電位差(膜電位)を保っている。これは神経伝達や筋収縮に必須だ。
【正常な膜電位】
細胞外:0 mV(基準)
細胞内:-50〜-90 mV(細胞内が負)
この電位差を作るのが:
- ATPアーゼ:イオンの交換
- チャネル:K⁺やNa⁺が選択的に外へ漏れる → 結果として細胞内が負に保たれる
もしHCO₃⁻(マイナス電荷)だけを外へ出し続けたらどうなるか?それは、皆さんもよく知るだろう有名なフグ毒、テトロドトキシンを思い浮かべるといいだろう。
【テトロドトキシンの作用】
1.Na⁺チャネルに結合
→ Na⁺の流入を完全に阻止
2.活動電位が発生できなくなる
神経細胞:興奮伝達が停止
筋細胞:収縮命令が届かない
3.全身の神経・筋肉が麻痺
- 感覚麻痺(しびれ)
- 運動麻痺(動けない)
- 呼吸筋麻痺(呼吸停止) 4.数時間で死亡
なぜこれほど致命的なのか? 膜電位の変化が、生命活動の根幹だからだ。
神経細胞が信号を伝える仕組みを見てみよう:
【神経の活動電位】
安静時:
細胞内 -70 mV(K⁺が多い)
細胞外 0 mV(Na⁺が多い)興奮時(刺激を受けた):
1.Na⁺チャネルが開く
2.Na⁺が細胞内に流入
3.膜電位が上昇:-70 mV → +30 mV
(脱分極)
4.この電位変化が隣の細胞に伝わる
→ 神経信号の伝達回復:
5.K⁺チャネルが開く
6.K⁺が細胞外に流出
7.膜電位が下降:+30 mV → -70 mV
(再分極)
8.NKAがNa⁺とK⁺を元の場所に戻す
テトロドトキシンは、このステップ1をブロックする。Na⁺チャネルが開かなければ、脱分極が起きない。神経信号が伝わらない。
結果:
- 感覚神経が動かない → 痛みを感じない、触覚がない
- 運動神経が動かない → 筋肉を動かせない
- 呼吸筋が動かない → 呼吸停止 → 死亡
つまり、膜電位の制御が狂うと、生物は死ぬ。よって、HCO₃⁻の排出には、電荷を補償する仕組みが絶対に必要なのだ。
さらに面倒なことにCAでCO₂を捕獲されるとき、HCO₃⁻を作ると同時に、H⁺(プロトン)も生成する。
CO₂ + H₂O → H₂CO₃ → H⁺ + HCO₃⁻
つまり、CO₂を処理すればするほど、細胞内は酸性に傾いていく。体液が酸性に傾き過ぎると、酵素の失活やたんぱく質の変性によって、代謝が止まってしまう。これをアシドーシスという。
ルブラリスでは、海水からのCO₂侵入を防ぐために、CAが超高濃度(地球の5〜8倍)で常時フル稼働している。その結果による急速なpH低下、HCO₃⁻の蓄積、膜電位の維持、これらすべてを同時に解決する必要がある。それでは、ルブラリスのカンブリア紀相当期の魚類は、この過酷な環境にどう適応したのか。
細胞内pHの適応
まず重要なのは、ルブラリスの魚類は細胞内pHそのものを海水に適応させたという点だ。地球の魚類の細胞内pHは7.2〜7.4だが、これは地球の海水pH(約8.1)に最適化された結果である。ルブラリスの海はpH 8.2〜8.5——当然、細胞内pHもそれに追随するべきだ。
しかし、完全には追随できない。なぜなら、生化学的制約があるからだ。代謝酵素の大半は、pH 7.0〜7.8で最適活性を示す。これはアミノ酸の解離定数や活性部位の電荷分布といった、化学的な基本制約だ。pH 8.0以上になると、酵素活性は50〜70%に低下し、DNA/RNAも加水分解が加速する。
進化的妥協点として、ルブラリスの魚類は細胞内pH 7.5〜7.7に落ち着いた。地球より0.3〜0.5高く、海水より0.5〜1.0低い。
細胞内(pH 7.6):[H⁺] = 2.5 × 10⁻⁸ M
海水(pH 8.4):[H⁺] = 4.0 × 10⁻⁹ M
勾配:約6倍
→ H⁺は濃度勾配に従って自然に細胞外へ流れる
→ 排出コストが大幅に削減される
ただし代償もある。酵素活性が85〜90%に低下するため、代謝効率が10〜15%落ちる。しかし、H⁺排出に大量のエネルギーを使い続けるより、遥かに効率的だ。
統合型鰓上皮細胞
第二の解決策は、細胞の統合だ。地球の魚類の鰓には、機能の異なる二種類の細胞がある。呼吸上皮細胞(厚さ1〜5 µm、ガス交換専門)と、先ほど述べた塩類細胞(厚さ20〜40 µm、浸透圧調節専門)だ。この分離には理由がある。ガス交換には「薄さ」が、浸透圧調節には「ポンプの高密度配置」が必要で、両者はトレードオフの関係にあるからだ。
しかしルブラリスでは、この分離が不要になった。理由は単純だ。ルブラリスの鰓細胞は、すでにCA超高発現(細胞質の5〜10%)や各種ポンプ・交換体で「厚く」なっている(10〜20 µm)。ミトコンドリア密度も高い(体積の30〜40%)。つまり、地球の「塩類細胞」にすでに近い構造なのだ。ここに浸透圧調節機能を統合しても、追加の厚み増加は10〜20%程度で済む。ガス交換効率への影響は限定的だ。
結果として、ルブラリスの鰓は細胞一種類で全機能を担う。
【細胞の機能】
- O₂取り込み(拡散)
- CO₂トラップ(CA超高発現)
- H⁺排出(三重システム)
- HCO₃⁻排出(Cl⁻/HCO₃⁻交換体)
- pH維持(フィードバック制御)
- 浸透圧調節(Na⁺-K⁺-ATPase)
すべてが一つの細胞で完結する。細胞を分離する必要がないため、同じミトコンドリアでATP生産、同じ膜で複数のポンプ・交換体を配置でき、効率が向上する。
細胞の動き:
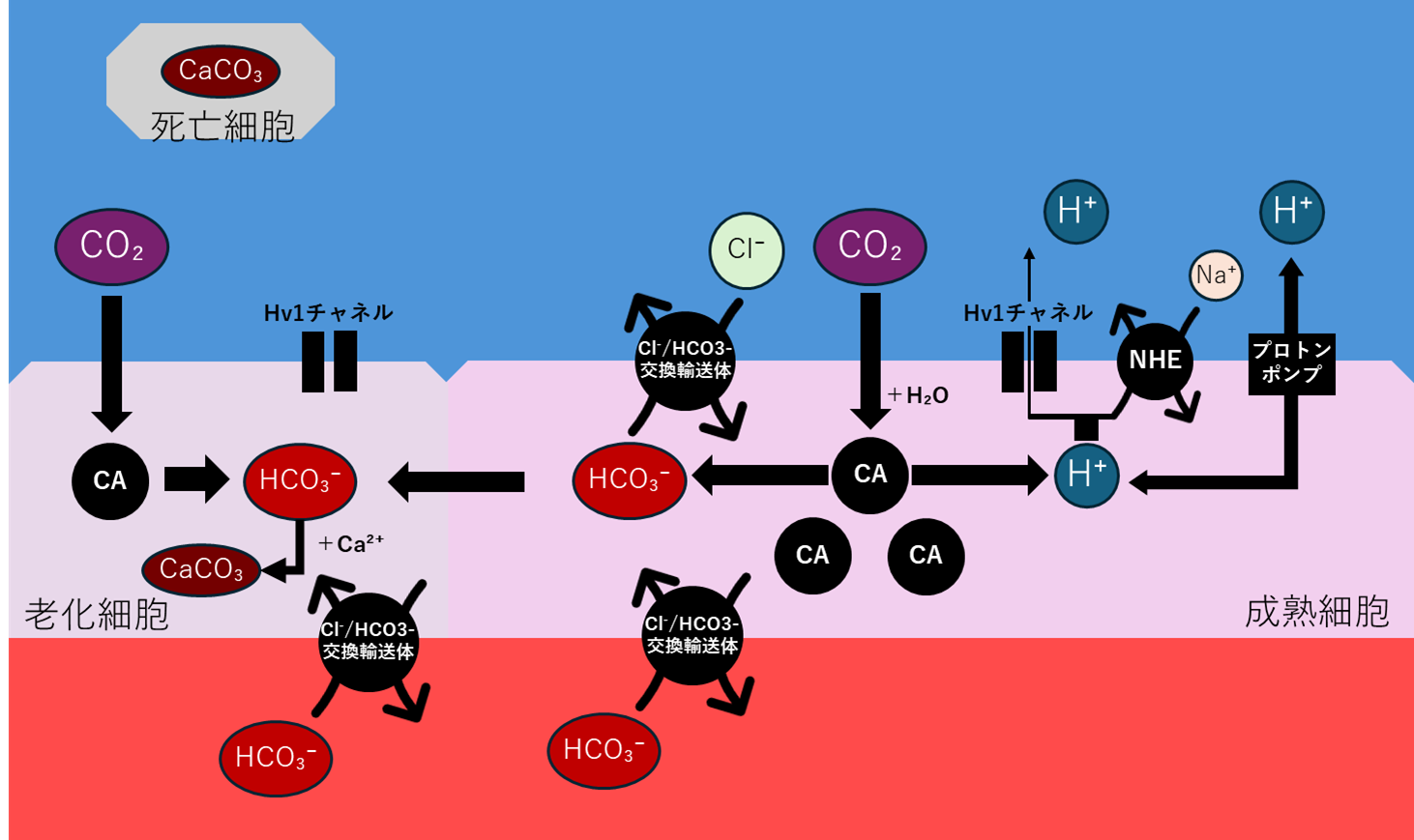
H⁺排出の三重防衛システム
先ほど述べたH⁺の排出問題については三層のシステムでこれに対応する。
重要なのは、海水側のpHは変化しないという点だ。ルブラリスの海水は、DIC濃度30〜50 mMという強力な炭酸緩衝系だ。鰓から排出されるH⁺など、海水にとっては微々たるもので、pHは常にpH 8.2〜8.5を保つ。
さらに、海水は常に流れている(1〜5 cm/秒)。わずかに酸性化した海水は、すぐに流され、新しいアルカリ海水が来る。つまり、鰓周辺の海水はほぼ完全に一定pHだ。よって、濃度勾配は常に維持される。
第一層:Hv1チャネル
最も重要なのが、電位依存性プロトンチャネル(Hv1)だ。これは4回膜貫通の単純なタンパク質(273アミノ酸)で、H⁺を選択的に透過させる。
Hv1の最大の利点は、ATP不要という点だ。細胞内pH 7.6と海水pH 8.4の濃度勾配に従って、H⁺は自然に細胞外へ流れる。物理法則に従うだけで、エネルギーを使わない。
ルブラリスの鰓細胞は、Hv1を地球の魚の3〜5倍も高密度で発現している。細胞あたり10⁵〜10⁶個、1個あたり毎秒10⁴〜10⁵個のH⁺を透過させる。これでH⁺排出の40〜60%を担う。
もう一つの利点は、構造が単純で酸化に強いことだ。システイン残基が少なく、老化してもほぼ機能を保持する。若い細胞から老化細胞まで、すべてが同等にH⁺排出に寄与できる。
第二層:H⁺-ATPase(精密制御)
しかしHv1だけでは不十分だ。チャネルは電位依存性で開閉するため、代謝の変動に即座には対応できない。精密な制御には、能動輸送が必要だ。
それが可能なのが、H⁺-ATPaseだ。このポンプは、1分子のATPを消費して、1個のH⁺を細胞外へ排出する。Hv1より遥かに低速だが、その代わり精密に制御できる。
H⁺生成 > Hv1排出
→ 細胞内pHが低下傾向(pH < 7.6)
→ H⁺-ATPaseが追加でH⁺を排出
→ pH 7.6に戻すH⁺生成 < Hv1排出
→ 細胞内pHが上昇傾向(pH > 7.6)
→ H⁺-ATPaseを減速/停止
→ CA反応が自動的にH⁺を生成(平衡シフト)
→ pH 7.6に戻す
H⁺-ATPaseは、細胞内pHを狭い範囲に保つフィードバック制御装置だ。H⁺排出の20〜30%を担う。
第三層:Na⁺/H⁺交換体(NHE、補助システム)
第三のシステムが、Na⁺/H⁺交換体(NHE)だ。これは、Na⁺(外→内)とH⁺(内→外)を1:1で交換する。駆動力は、Na⁺-K⁺-ATPase(NKA)が作るNa⁺勾配で、海水Na⁺の470 mM、細胞内Na⁺の10〜15 mMという勾配を利用して、H⁺を排出する。ATP不要の「二次性能動輸送」だ。
NHEの役割はpHと浸透圧の統合制御だ。H⁺を出しながらNa⁺を取り込むため、pHと電解質バランスを同時に調整できる。H⁺排出の10〜20%を担う。ただし、ルブラリスではNa⁺過剰が問題になる(後述)ため、NHEは控えめに使われる。主力はあくまでHv1だ。
炭酸塩排出とその代償
CA反応で生成したHCO₃⁻を処理するのが、Cl⁻/HCO₃⁻交換体だ。地球の塩類細胞にもあった、あの交換体だ。しかしルブラリスでは、その重要性が桁違いに高まる。
この交換体は、細胞膜を貫通するタンパク質で、陰イオン同士を1:1で交換する。鰓上皮細胞には、この交換体が血液側と海水側の二箇所に配置されている
この二つの交換体が協調して働くことで、血液中のHCO₃⁻を海水へ排出する。Cl⁻を取り込むことで、電荷バランスが保たれることにより、前述の「陰イオンだけを出し続けると膜電位が崩れる」問題も解決される。
ここで、地球との決定的な違いが浮上する。ルブラリスでは、CAの逆反応が不要なのだ。地球の魚類を思い出してほしい。組織で生成したCO₂を赤血球内でHCO₃⁻に変換し、血液で運び、鰓で再びCO₂に戻してから海水へ放出する。鰓はCO₂ガスしか排出できないため、血液中のHCO₃⁻を一旦CO₂に戻さなければならない。この逆反応のため、赤血球にも鰓の毛細血管にも高濃度のCAが必要だ。
しかしルブラリスは違う。代謝したCO₂は赤血球でHCO₃⁻したら戻す必要がない。 鰓がHCO₃⁻として直接海水へ排出できるからだ。海水はpH 8.2〜8.5という弱アルカリ性で、HCO₃⁻はむしろ安定な化学形態だ(海水DIC 30〜50 mMの大半がHCO₃⁻形態)。わざわざCO₂に戻す必要がない。この一方向性がもたらす利点は三つある。
1.赤血球CAの削減
地球:双方向必要 → 高濃度CA
ルブラリス:一方向のみ → 地球の50〜70%で十分2.鰓CAの局在化
地球:鰓の基底側(血液側)にもCA必要(逆反応のため)
ルブラリス:頂端側(海水側)のみCA超高発現理由:
- 頂端側:海水侵入CO₂を即座にHCO₃⁻固定(超高濃度必須)
- 基底側:血液からのHCO₃⁻をそのまま受け取る(CA不要)
3.熱力学的効率
順反応(pH 7.6):ΔG ≈ −5 kJ/mol(やや有利)
逆反応(pH 7.6):ΔG ≈ +5 kJ/mol(やや不利)
→ 逆反応を避けることで、エネルギー損失を削減
しかし、その過程でCl⁻が血液に蓄積する。これは避けられない副作用だ。なぜCl⁻なのか? 理由は二つある。第一に、HCO₃⁻は陰イオンなので、電荷を補償するために別の陰イオンを入れなければ膜電位が崩れる。第二に、海水に豊富にある陰イオンはCl⁻しかない(海水Cl⁻:550 mM)。さらに、電気的中性を保つため、Na⁺も追随して上昇する。
この高塩血症は、浸透圧を上昇させる。放置すれば、細胞から水が流出し、脱水に至る。ルブラリスの魚は、この過剰な塩を排出しなければならない。
しかし、ここでさらなる問題が浮上する。食事だ。ルブラリスの魚は、浅海のプランクトンや小魚を食べる。彼らの体液も高塩(Na⁺、Cl⁻が豊富)だ。つまり、食事からも大量のNa⁺、Cl⁻が入ってくる。
したがって、地球の肝門脈のような血液構造になるだろう。地球の動物では、腸管から吸収された栄養素は、まず肝門脈を経由して肝臓へ送られる。肝臓で毒素を解毒し、栄養素を処理してから、循環系に入る。これは一種の「前処理システム」だ。
鰓を通した血液はまず塩類腺を通る。これにより、細胞が過剰な塩分をNaClとして排出してしまう。生物を学んだことがある人はなぜ腎臓ではないのかと思った方がいるかもしれない。それは、腎臓は本来、濾過と再吸収のシステムだからだ。
腎臓はネフロンという単位の集まりだ。
【ネフロンの構造】
糸球体(濾過装置):
血液 → 濾過 → 原尿
- 水、塩、尿素、グルコース、アミノ酸など
- 小分子はほぼすべて濾過される
- 血球やタンパク質は濾過されない ↓ 尿細管(再吸収装置): 原尿 → 再吸収 → 血液
- 必要な物質を取り戻す
- 水:90〜99%再吸収
- Na⁺:99%再吸収
- グルコース:100%再吸収
- 尿素:一部再吸収(濃度維持) ↓ 残り → 尿として排出
【腎臓での塩排出】
- 糸球体で血液を濾過
- 塩も水も一緒に出る
- 尿細管で水を再吸収
- 塩を濃縮する
- しかし濃縮限界がある
このシステムの問題は、塩を排出するために大量の水を使うことだ。濃縮限界は、浸透圧勾配で決まる。尿細管の細胞は、能動輸送でNa⁺を尿中に出すが、その駆動力(ATP)には限界がある。ある程度濃縮すると、浸透圧差が大きくなりすぎて、それ以上濃縮できない。そのため、大量の水に溶かすしか方法がない。しかし、その方法でいくと脱水で死んでしまう。
腎臓は万能ではない。濾過・再吸収という仕組みは、通常の老廃物(尿素など)の排出には優れているが、大量の塩を効率的に排出するには向いていない。だから、魚は鰓で塩分を輩出しているのだ。
しかし、地球には、この問題を解決した動物がいる。海鳥と海爬虫類だ。彼らは海水を飲む。海水の塩濃度(約550 mM NaCl)は、彼らの体液(約150 mM)より遥かに高い。腎臓だけでは処理できない。そこで進化したのが塩類腺だ。
塩類腺は、Na⁺-K⁺-ATPaseを超高密度で発現した細胞の集まりだ。
【塩類腺細胞の働き】
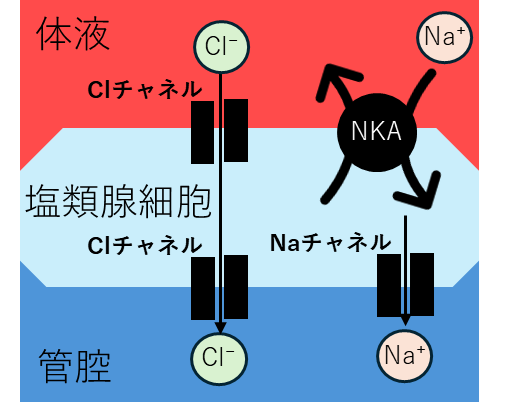
- 血液からNa⁺を取り込む(基底膜) Na⁺-K⁺-ATPase:Na⁺(血液→細胞)
- 細胞内からNa⁺を分泌(頂端膜) Na⁺チャネル:Na⁺(細胞→管腔)
- Cl⁻が電気的に追随 Cl⁻チャネル:Cl⁻(細胞→管腔)
- 浸透圧で水が追随 水チャネル:H₂O(細胞→管腔) 結果: 高濃度NaCl溶液(1000〜1200 mM)が管腔に蓄積
これの排出口は、鰓蓋の後縁付近、鰓孔のすぐ脇にある。泳いでいると、排出された塩溶液は海水流に即座に希釈される。
細胞の階層的役割分担
ルブラリスの鰓上皮細胞は、成熟度によって役割が分かれる。細胞のターンオーバー周期は5〜10日(平均7日)程度だ。地球の鰓(7〜14日)よりやや速いが、消化器粘膜(3〜5日)よりは遥かに遅い。
若い細胞(0〜2日目)
基底膜近くの幹細胞から分裂した若い細胞は、まだCAや各種ポンプの発現が不十分だ。しかしHv1は早い段階から機能する。この段階では、主にH⁺排出に寄与し、本格的な機能は発揮していない。
成熟細胞(2〜8日目)
成熟細胞が、鰓の主力だ。CA発現が最大(細胞質の5〜10%、ただし頂端側に局在)に達し、Cl⁻/HCO₃⁻交換体、H⁺-ATPase、NHE、Na⁺-K⁺-ATPaseもすべて高密度で発現している。
CO₂トラップ、H⁺排出、HCO₃⁻排出、pH管理、浸透圧調節——すべてを高効率で実行する。成熟細胞が、鰓全体の機能の70〜80%を担う。
老化細胞(8〜10日目)
老化が進むと、CA活性が低下する(30〜50%に)。Cl⁻/HCO₃⁻交換体の効率も落ちる。しかしHv1は、構造が単純なため、ほぼ正常に機能し続ける。
ここで何が起きるか? 隣接する成熟細胞から、HCO₃⁻がわずかに漏れ出る。細胞間隙を通じて、老化細胞へ流入する。老化細胞は、HCO₃⁻を処理する能力が低いため、HCO₃⁻が蓄積していく。
細胞内HCO₃⁻濃度が20〜30 mMに達すると、血液由来のCa²⁺(通常0.0001 mM、老化時0.001〜0.01 mM)と結合し始める
HCO₃⁻ + Ca²⁺ → CaHCO₃⁺(可溶性)
2HCO₃⁻ + Ca²⁺ → CaCO₃↓ + H₂O + CO₂
→ 微小結晶(<1 µm)の形成
CaCO₃微小結晶は、小胞体やリソソームに蓄積する。HCO₃⁻過剰でpHが上昇(pH 7.8〜8.0)、Ca²⁺過剰でカスパーゼが活性化、結晶の機械的ストレス——これらが複合して、アポトーシス(プログラム細胞死)が誘導される。細胞は縮小し、隣接細胞が隙間を埋める。そして老化細胞は、HCO₃⁻、Ca²⁺、微小結晶を全部含んだまま、海水へ排出される。
これは一種の「分泌」だ。老化細胞を、炭酸塩とカルシウムの「ゴミ袋」として使う。地球の腎臓でも、尿中のCa²⁺過剰時に集合管細胞が結晶を蓄積してアポトーシスで排出する例がある。ルブラリスの鰓は、この戦略を日常的に使っているのだ。
酸素の取り込み
ここまでCO₂の話ばかりしてきたが、もちろん鰓の本来の役割は酸素の取り込みだ。しかし、この点に関しては、ルブラリスは地球と大差ない。
大気O₂濃度はルブラリス5%、地球21%と大きく異なるが、全圧がルブラリス4.5気圧、地球1気圧のため、分圧(ppO₂)はほぼ同じになる。溶存酸素量も同等だ。なので、地球と同様に、酸素の取り込みは、単純な拡散で行われる。
地球と異なり、ルブラリスの鰓細胞はやや厚い(10〜20 µm vs 地球の1〜5 µm)。しかしミトコンドリア密度が高く(30〜40%)、細胞内での酸素拡散距離が短いため、問題にならない。
さらに、血液中の酸素運搬も地球と同じヘモグロビン系で行われる。唯一の違いは、血液pHがやや高い(pH 7.6〜7.8 vs 地球の7.4)ため、ヘモグロビンの酸素親和性がわずかに変わることだが、これも進化的に最適化される。
ここで、決定的な制約が浮上する。ルブラリスの魚は、止まれないのだ。
地球の魚類も鰓換水は必須だが、多くは止まれる。底生魚(ヒラメ、カレイ、アンコウ)は海底でじっとしながら、口と鰓蓋を交互に動かす「頬ポンプ」で換水する。停止時間は数分〜数十分、場合によっては数時間も耐えられる。
一部の高速遊泳魚(マグロ、サメの一部)だけが、口を開けて泳ぐことで受動的に換水する「ラム換気」を使う。彼らは止まると窒息するが、これは地球では特殊適応だ。
しかしルブラリスでは、ラム換気が標準になる。
理由は、海水の緩衝能の強さだ。鰓を通過した海水は、わずかにCO₂が増え、わずかにO₂が減る:
【鰓通過による変化】
pCO₂:150 mmHg → 155〜160 mmHg(+5〜10 mmHg)
ppO₂:0.225 atm → 0.10〜0.15 atm(約50%消費)
pH:8.30 → 8.29(ほぼ変化なし、緩衝能のため)
pHの変化は測定誤差レベルだが、pCO₂とppO₂は確実に変わる。もし魚が止まると、鰓周辺の海水が「使い古される」。局所的にpCO₂が170〜180 mmHgまで上昇し、ppO₂が0.05 atm以下に低下する。
血液pCO₂(50〜100 mmHg)との逆勾配がさらに悪化し、CO₂排出がほぼ不可能になる。酸素も不足する。
結果
停止時間と症状:
0〜30秒:正常
30秒〜1分:血液pCO₂上昇、息苦しさ
1〜3分:呼吸性アシドーシス、意識混濁
3〜5分:筋力低下、活動不能
5分以上:窒息の危険
ルブラリスの魚は、地球の魚より遥かに短時間しか止まれない。頬ポンプを動かし続けることもできるが、そのエネルギーコストは基礎代謝の3〜5%にもなる。すでに高コストの呼吸系に、さらに上乗せされる。
より効率的なのは、泳いで水流を作ることだ。ラム換気なら、頬ポンプ不要で、遊泳コストは低速巡航時で1〜2%に収まる。正味2〜4%の節約だ。よって、ルブラリスではラム換気が主流になる。そして、ラム換気の魚は止まれない。睡眠時も泳ぎ続けるか(半球睡眠、地球のイルカ型)、強い水流の中で定位する(口を開けてじっとしている)必要がある。ただし水流利用も、数分〜数十分が限界だ。
この「動き続けなければならない」という制約が、生態を大きく変える。底生生活は放棄され、浅海の遊泳性が主流になる。そして、常に泳ぐためには、浮き袋が決定的に重要になる。浮力調整ができなければ、沈まないために常に鰭を動かし続けねばならず、エネルギーコストがさらに3〜5%増えるからだ。
最後に、これらすべてのシステムを動かす実際のコストを計算してみよう。
- 鰓:
- CA維持:0.3〜0.7%
- H⁺排出:1〜2%
- Cl⁻/HCO₃⁻交換:2〜3%
- Na⁺-K⁺-ATPase:1〜2%
- ターンオーバー:0.3% 小計:4.6〜8.0%
- 塩類腺(鰓後処理):2.5〜4%
- ラム換気:1〜2%
総計:8.1〜14.0%
地球の魚類:4〜7%
差:約2倍
鰓系の運転コストは基礎代謝の9〜14%(状況により最大17%)で高い。しかし、初期器官として十分に現実的だ。地球でも、初期の複雑器官は高コストから始まった。カンブリア紀の複眼は5〜10%、初期の翼(始祖鳥)は飛行時20〜40%を食い、常時稼働系の心臓は5〜10%、脳は人間で20%(魚類2〜5%)を占める。それでも進化は成立した。
重要なのは相対性能だ。ルブラリスのカンブリア相当期には誰も「効率的な鰓」を持たない。呼吸コスト20%の群より15%の群が、15%より10%が、順に競争を制す。絶対値ではなく、差分が運命を分ける。
総エネルギー配分
- 基礎代謝(心臓・脳・肝など):40〜50%
- 呼吸・排泄・換気:9〜14%(上限17%)
- 消化・吸収:5〜10%
- 遊泳(ラム換気の運動費用):上記に一部内包
- 成長・繁殖:10〜18%
- 予備(ストレス対応など):5〜15% 合計:100%
成長・繁殖に回せる配分は、地球魚(15〜25%)より5〜10%低い。これは制約だが致命的ではない。代償としてK戦略(少産・長寿・高親投資)へシフトすれば十分に整合する。
この「高いが致命的でない」コストが、進化の物語を駆動する。生存が可能だから多様化し、重いコストが効率化圧を恒常的に与える。その圧力が、浮力制御系の価値を極限まで引き上げた。
ここから地上呼吸まで至る呼吸器進化に移るはずであったが、この時点であまりに長くなり過ぎたので、一旦の区切りにしたい。
おまけ:
1.原始生物の誕生
生命はどこで生まれたのか?深海の熱水噴出孔や火山周りの間欠泉、天然原子炉など諸説あるが、現在もっとも有力視される仮説の一つが、アルカリ性の深海熱水噴出孔だ。そこには生命誕生に必要な三つの条件が揃っている。
- エネルギー源:熱水と海水の温度差、化学物質の酸化還元反応
- 化学物質:H₂、CO₂、H₂S、Fe²⁺など、有機物合成の材料
- 濃度勾配:噴出孔と海水の間でpH、温度、化学濃度の勾配が自然に形成される
特に重要なのが最後の「濃度勾配」だ。生命とは本質的に「勾配を利用して秩序を作り出すシステム」である。熱水噴出孔の壁面では、酸性の熱水(pH 3〜5)とアルカリ性の海水(pH 8〜9)が接触し、自然に多孔質の鉱物壁(硫化鉄など)が形成される。この壁の無数の微小な孔が、最初の「細胞膜」の役割を果たしたと考えられている。
地球での生命誕生(40〜38億年前):
地球の初期海洋は、前回述べたような超酸性雨の時代を経て、やや酸性〜中性(pH 6〜7)に落ち着いていた。大気は依然として高CO₂・低O₂で、紫外線はオゾン層がないため地表まで到達していた。だが深海の熱水噴出孔は、紫外線から守られた理想的な「ゆりかご」だった。
最初の代謝は極めてシンプルだった:
H₂ + CO₂ → 有機物 + H₂O
水素ガス(H₂)を電子源として、二酸化炭素(CO₂)を炭素源とする。これは「化学合成」と呼ばれ、光も酸素も必要としない。現在でも深海の熱水噴出孔周辺には、この代謝だけで生きる古細菌(アーキア)が存在する。彼らはおそらく、地球最古の生命の直系の子孫だ。
特に重要なのがメタン生成菌だ:
CO₂ + 4H₂ → CH₄ + 2H₂O
この反応は発熱反応で、エネルギーを得られる。生成したエネルギーを使ってATP(生命のエネルギー通貨)を作り、細胞を維持・増殖させる。CO₂が豊富な環境では、この代謝は極めて有利だった。
ルブラリスでの生命誕生(40億年前):
ルブラリスの場合、状況はさらに劇的だった。前回で述べたように、全球凍結とその後の超酸性雨により、海洋は高DIC(溶存無機炭素)・高アルカリという極端な環境になっていた。
全球凍結が解凍されてから数百万年後、海洋の化学組成は以下のようになっていたとしよう。:
- pH:9.0〜9.5(地球の初期海洋は6〜7)
- DIC濃度:数十〜数百 mM(地球の10〜50倍)
- CO₂分圧(海水中):0.2〜0.3 atm相当(地球の2〜3倍)
- Mn²⁺濃度:数百 nM〜μM(地球の10〜100倍)
この環境は、一見すると生命にとって過酷に見える。pH 9.5という強アルカリ性は、現代の地球生物の大半の細胞を殺す。しかし深海熱水噴出孔では、熱水自体は依然として酸性(pH 3〜5)だ。つまり、酸性の熱水とアルカリ性の海水が接触する——この強い勾配こそが、生命誕生の鍵となった。
さらに決定的だったのが、CO₂の豊富さだ。地球の初期海洋でも十分CO₂は豊富だったが、ルブラリスはその比ではない。DIC濃度が10〜50倍ということは、生命にとって「炭素源を探す必要がない」ということだ。
最初の代謝は地球と同じく:
H₂ + CO₂ → 有機物 + H₂O
しかし反応速度が桁違いに速かった。律速段階(反応全体で最も遅い段階)は通常、CO₂の取り込みだ。ところがルブラリスでは、CO₂が過剰に存在するため、この制約がほぼない。結果として、メタン生成菌相当の生物は、地球の同類より数倍速く増殖できた可能性がある。
しかし、ルブラリスの生命には地球にはない課題があった。高アルカリ環境への適応だ。
pH 9〜9.5という環境では、次の三点が致命的になる。
1. 膜が壊れやすい
地球の細菌や真核生物は「エステル脂質」で膜を作る。しかし、高校化学で習った方は理解できるだろうが、エステルは塩基(OH-)によって加水分解される(けん化)。これにより膜脂質が切断されて、透過性が上がり、イオン勾配や膜電位が保てない。つまり、細胞膜が穴だらけになるということだ。
2. 核酸・酵素が働きにくくなる
RNA はアルカリ条件で 2'-OH による自己切断(アルカリ性 RNA 分解)が進む。また、タンパク質は水素結合が脱プロトン化(H⁺が外れる)によって水素結合・塩橋が崩れ、伸びたバネが縮むように形が変わる。そうなると、機能を失ってしまう(失活)。
3. エネルギーの“落差”が作れない
多くの生物は、膜の内外にプロトン勾配(pH差)と膜電位を作り、その化学浸透(プロトン駆動力)で ATP 合成酵素(F₁F₀-ATPase)が ATP(エネルギー通貨)を作る。外が強アルカリだと外側の H⁺ が少なく勾配が作りにくいうえ、劣化した膜からの漏れで維持も難しい。
よって、ルブラリスの最初の生命は、以下の適応を獲得した:
(1)エーテル脂質膜の採用
地球の細菌や真核生物は「エステル脂質」で膜を作るが、古細菌の一部、特にアルカリ環境に住む種は「エーテル脂質」を使う。エーテル結合(C-O-C)はエステル結合(C-O-CO-C)より加水分解に強く、高pHでも安定だ。
ルブラリスの生命は当初から、エーテル脂質を主体とする膜を持っていた可能性が高い。これは偶然ではなく、高pH環境での自然選択の結果だ。エステル脂質の膜を持つ個体は、数世代で膜が崩壊して死滅したはずだ。
(2)HCO₃⁻/CO₂変換酵素の発達
pH 9.5の海水では、CO₂の大半が炭酸水素イオン(HCO₃⁻)や炭酸イオン(CO₃²⁻)として存在する。
化学平衡の計算では:
pH 9.5の場合:
CO₂:約0.1%
HCO₃⁻:約90%
CO₃²⁻:約10%
つまり、CO₂そのものは意外と少ない。しかし総DIC量が多いため、絶対量では地球より豊富——これが重要だ。
生命はHCO₃⁻を直接利用できない。代謝で使えるのはCO₂だけだ。
そこで炭酸脱水酵素(CA:Carbonic Anhydrase)という酵素が重要になる:
HCO₃⁻ + H⁺ ⇄ CO₂ + H₂O
この酵素は地球の生物も持っているが、ルブラリスの生命ではさらに高活性・高濃度で発達した。細胞内のpHをわずかに下げて(pH 8程度)、HCO₃⁻からCO₂を遊離させる——このシステムが、初期の段階から確立されていた。
(3)プロトンポンプの強化
細胞内のpHを制御するため、H⁺-ATPaseという酵素が不可欠だ。これは膜に埋め込まれた酵素で、ATPを消費して細胞内からH⁺を汲み出すポンプだ。地球の好アルカリ性細菌も同様のシステムを持つが、ルブラリスの生命ではこれが「標準装備」として、より強力に発達していた。
興味深いのは、このプロトンポンプが後の進化——特に呼吸器官の進化——で決定的な役割を果たすことだ。これについては後述する。
(4)チャネルによる微調整
チャネルは膜を貫くたんぱく質の細孔で、ATPを使わずにイオンが濃度差や電位差(電気化学勾配)に従って通過する。ポンプが「能動的に汲む」道具だとすれば、チャネルは「栓を開け閉めして流れを調節する」道具である。ルブラリスではH⁺の漏れチャネルは抑制し、電位依存性・リガンド依存性・機械受容性などのゲート付きチャネルを必要な時だけ開く運用で、膜電位と浸透圧を素早く・無駄なく微調整する。
では、ルブラリスでの生命誕生は地球より早かったのか、遅かったのか?
答えは、惑星形成からの経過時間で見れば、地球とほぼ同時期だ。
なぜ高DIC・高pHという極端な環境でも、地球と同じ時間スケールで生命が誕生できたのか? 理由は二つある。
- CO₂の豊富さ:律速段階が解消され、代謝効率が高い
- 勾配の強さ:pH 3〜5の熱水とpH 9.5の海水——地球(pH 3〜5 vs pH 6〜7)より大きな勾配
特に(2)は重要だ。より大きなpH勾配は、より強い化学ポテンシャル(自由エネルギー)を意味する。これを利用できる代謝系が偶然生まれれば、爆発的に増殖できる。自然選択の圧力も強まり、進化が加速する可能性がある。
こうして、ルブラリスの海にも生命が誕生した。地球と同じくメタン生成菌・酢酸生成菌に相当する化学合成微生物が最初の住人となった。ただし彼らの海は、pH 9.5、DIC数百mM、Mn²⁺が豊富という高アルカリ・高炭酸の世界であり、その出発点に適応して膜化学(エーテル脂質)とエネルギー獲得機構が地球とは異なる方向へ最初から振られている。
2. 光合成生物の登場
化学合成だけで生きる生命は、熱水噴出孔という限られた「オアシス」に縛られていた。H₂やH₂Sといった還元物質は、火山活動のある場所でしか得られない。これでは生息域を広げられない。
しかし海には、無尽蔵のエネルギー源があった。光だ。
主星セファエルは暗いとはいえ、海面には依然として光が降り注いでいた。地球の海面照度の約25%——曇天の地球と同程度だ。この光エネルギーを使えれば、化学物質に頼らず、海洋全体に生息域を広げられる。そこに至る進化圧は強烈だった。
地球での光合成進化:
まず地球での経緯を整理しよう。光合成は一度に完成したのではなく、段階的に進化した。
段階1:嫌気的光合成(35億年前〜)
最初の光合成生物は、硫黄細菌(紫色硫黄細菌、緑色硫黄細菌)だった。彼らは硫化水素(H₂S)を電子源として使った:
2H₂S + CO₂ + 光 → (CH₂O) + H₂O + 2S
H₂Sから電子を奪い、その電子でCO₂を還元して有機物(糖)を作る。副産物として硫黄(S)が出る。酸素(O₂)は出ない——これが「嫌気的」光合成と呼ばれる理由だ。
使う色素は「バクテリオクロロフィル」。クロロフィル(葉緑素)の仲間だが、吸収波長が異なる。バクテリオクロロフィルは近赤外(700〜1000 nm)を吸収し、可視光はあまり使わない。そのため紫色硫黄細菌は紫〜赤紫色、緑色硫黄細菌は暗褐色に見える。
この光合成は画期的だったが、限界もあった。H₂Sがある場所でしか生きられない。熱水噴出孔の周辺や、火山性の湖沼に限定される。海洋全体には広がれなかった。
段階2:酸素発生光合成(27億年前〜)
そこに革命が起きた。シアノバクテリア(藍藻)の出現だ。彼らは水(H₂O)を電子源として使い始めた:
2H₂O + 光 → O₂ + 4H⁺ + 4e⁻
CO₂ + 4H⁺ + 4e⁻ → (CH₂O) + H₂O
────────────────────────
全体:2H₂O + CO₂ + 光 → (CH₂O) + H₂O + O₂
水は地球上どこにでもある。これで生息域の制約が消えた。シアノバクテリアは爆発的に広がり、海洋全体を覆った。副産物として出る酸素(O₂)が、大気に蓄積し始めた——これが後の「大酸化イベント」(24億年前)につながる。
しかし水分解は容易ではない。水はH₂Sより遥かに安定で、電子を奪うには高い酸化電位が必要だ:
H₂S → S + 2H⁺ + 2e⁻ (酸化電位:−0.27 V)
H₂O → ½O₂ + 2H⁺ + 2e⁻ (酸化電位:+0.82 V)
1ボルト以上の差がある。つまり、水分解には強力な「酸化剤」が要る。シアノバクテリアはこれを、Mn₄CaO₅クラスターという金属錯体で実現した。4つのマンガン(Mn)イオンと1つのカルシウム(Ca)イオン、5つの酸素(O)が組み合わさった、極めて複雑な触媒だ。
この触媒の設計図(タンパク質構造)を、進化で獲得するのに数億年かかった。27億年前の酸素発生光合成の登場は、約8億年の試行錯誤の末の「大発明」だったのだ。
シアノバクテリアが使う色素は「クロロフィルa」。これは可視光の青(430 nm)と赤(660 nm)を吸収する。緑色(500〜600 nm)はほとんど吸収しないため、反射される——これが植物が緑色に見える理由だ。
なぜ緑を使わないのか? これは太陽のスペクトルと関係がある。太陽光は緑色付近(500〜550 nm)にピークがあり、光量が最も多い。一見、ここを使えば効率的に思える。しかし実際には、青と赤を使った方が、化学反応に必要な「高エネルギー光子」を効率よく集められる。緑色は中途半端なエネルギーで、使いにくいのだ。
進化的には、初期の光合成色素(バクテリオクロロフィル)が近赤外を使っていたため、シアノバクテリアが可視光に進出する際、既に近赤外を使う他の細菌との競合を避けるため、青と赤にシフトした——という説が有力だ。
ルブラリスでの光合成進化:
ルブラリスでも、基本の段階は同じだが、光の色(近赤外優勢)と海の化学(高アルカリ・高DIC・Mn豊富)が、地球とは違う枝分かれを生んだ。
段階1:嫌気的光合成(35億年前〜)
最初は地球と同じく、H₂Sを使う硫黄細菌相当の生物が現れた:
2H₂S + CO₂ + 光 → (CH₂O) + H₂O + 2S
使う色素はバクテリオクロロフィル系だが、その吸収波長の最適点は、主星セファエルのスペクトルと海水の吸収特性で決まる。
主星セファエルの放射スペクトルを思い出そう:
放射ピーク:920 nm(近赤外)
可視光(400〜700 nm):全放射の約40%
近赤外(700〜1000 nm):全放射の約60%
つまり、入射エネルギーは近赤外に厚い。一方で、海水は波長が900 nmを超えると急激に光を吸収する。結果として、ごく浅い層(〜数m)に棲む群体では、主吸収を800–880 nm帯に置き、900 nm近傍は補助的に拾うという設計が進化的に有利になる。
このためルブラリスの硫黄細菌は、地球の紫色硫黄細菌(~800–850 nm)よりやや赤側にシフトした800–880 nm中心のバクテリオクロロフィル変異体を主力とし、環境次第で~900 nm帯を補助吸収する型も現れた。見た目は暗褐〜黒紫。可視域の反射が多く、近赤外を強く吸う「黒潮」として浅海底や湧水境界を覆う。
段階2:Mn触媒の獲得——地球より8億年早い革命
ここでルブラリス特有の分岐が起きる。酸素発生型の光合成が、地球(約27億年前)よりおよそ8億年早い時点(約35億年前)で立ち上がった、という仮説だ。鍵は海洋化学である。
地球の障壁:水分解の困難さ
地球の海はpH 7〜8の中性〜弱アルカリ性だった。この環境で水を分解する反応は:
2H₂O → O₂ + 4H⁺ + 4e⁻
酸化電位:+0.82 V(pH 7)
+0.82ボルトという高い電位が必要だ。
これを実現するには、Mn₄CaO₅という複雑な金属錯体を「発明」しなければならなかった。タンパク質の変異と自然選択を繰り返し、数億年かけて偶然この構造に到達した——それが地球の歴史だ。
ルブラリスの近道:OH⁻酸化
ところがルブラリスの海は、pH 9〜10の強アルカリ性だった。この環境では、水分子(H₂O)より水酸化物イオン(OH⁻)の方が遥かに多い。
化学平衡から:
pH 9の場合:
[H₂O]:55 M(ほぼ純水)
[OH⁻]:10⁻⁵ M
濃度比:OH⁻はH₂Oの約0.00002%
しかし反応性では:
OH⁻は既に部分的に電離している → 電子を奪いやすい
酸素発生反応(OER)の平衡電位は Nernst 則で
𝐸≈1.23−0.059×pH
と表せる。したがって、
- pH 7:𝐸≈+0.82V
- pH 9:𝐸≈+0.70 V
- pH 10:E≈+0.64 V
すなわち、中性に比べ 約 0.12〜0.18 V 低下 し、必要な酸化力のハードルがわずかに下がる(熱力学的優位)。
ここで、ある革命がおきる。活発な噴火で上空へ舞い上がった火山灰は、成層圏で紫外線と酸化的雰囲気にさらされ、灰表面の Mn(II) が Mn(III/IV) 酸化物(δ-MnO₂ など) としてナノ〜サブミクロンの皮膜・微粒子を形成する。粗粒は数日〜数週間で沈降し、微粒は半年〜数年滞留して広域に拡散、やがて海へ降灰する。大陸棚の浅海(降灰の収束帯)や海底火山近傍の湧昇域では、とくに Mn 酸化物微粒子のフラックスが高い。
同じ頃、海底火山の熱水プルームは嫌気的光合成アーキアを表層〜浅海へ運ぶ。乱流混合の中で細胞と Mn 酸化物(および灰片)は高頻度で衝突し、Ca²⁺/Mg²⁺ による電荷架橋や細胞外ポリマー(EPS)の粘着で複合凝集体をつくる。こうして 「アーキア+Mn 酸化物」の薄膜が浅海の上光層に点在する黒いマットとして張り付く。
利得は二重だ。第一に、Mn 酸化物が近赤外光下で電子の“足場”を提供し、OH⁻ の酸化(O₂ 発生)を助ける外付け触媒として働く(熱力学的ハードル低下に加え、速度論的ボトルネックの緩和)。第二に、灰と Mn 酸化物は可視〜UV を散乱し、無オゾン期の強い紫外からマット最上皮を遮蔽する。結果として、主星ピーク帯の近赤外に最適化したバクテリオクロロフィル群が活きる光環境が自然に整う。
この“外付け”段階で優位を得た系統では、やがて Mn 結合モチーフをもつ表層タンパクが増え、外付け粒子の恒常的保持 → 膜近傍への取り込みへと進化する。さらに Mn が 2〜4 個集まる簡素なクラスターを膜タンパク複合体内に固定し、反応中心の内蔵化が進む。こうして、近赤外吸収(850〜950 nm)を主とする色素系と、低電位側へシフトした OER 条件に合わせた Mn クラスターが結びつき、わずか数千万年〜1億年で Mn 触媒光合成が成立した(地球より圧倒的に速い)。
こうして誕生した酸素発生光合成生物を、地球の「シアノバクテリア(藍藻)」に相当する「メラノバクテリア(炭藻)」を持つ生物と呼ぼう。
メラノバクテリアの特徴:
- 色素:バクテリオクロロフィル誘導体
- 吸収波長:850〜950 nm(近赤外・主星スペクトルのピーク域)
- 補助色素:カロテノイド(赤〜黄色、可視光も少し吸収)
- 反応中心:Mn₂〜Mn₄錯体(OH⁻酸化触媒)
- 電子伝達系:地球の光化学系Iに類似(推定)
- 見た目の色: > - 主色素が近赤外吸収 → 可視光は大半が反射・透過 > - 補助色素が赤〜黄吸収 → 青〜緑がわずかに反射 > - 結果:黒褐色〜暗紫色
地球の植物は緑色だが、ルブラリスの植物はほぼ黒い。これは主星のスペクトルに最適化した結果だ。
光合成反応のまとめ
【光化学反応】
4OH⁻ + 光(近赤外850〜950 nm) → O₂ + 2H₂O + 4e⁻
(Mn触媒、酸化電位+0.3〜0.4 V)
【カルビン回路】
CO₂ + 還元力(NADPH) → (CH₂O)
【全体反応】
4OH⁻ + CO₂ + 光 → (CH₂O) + O₂ + H₂O
OH⁻が基質であることを除けば、地球の光合成と本質は同じだ。しかしこの小さな違いが、進化速度を劇的に変えた。
メラノバクテリアは爆発的に広がった。海洋表層を覆い尽くし、酸素を放出し続けた。やがて海水中の酸素濃度が上がり始めた。しかし、、当初は大気 O₂ が増えない。理由は二つある。
(1)鉄との反応(鉄沈殿)
海水中には鉄イオン(Fe²⁺)が大量に溶けていた。火山活動からの供給だ。酸素はまずこれと反応した:
4Fe²⁺ + O₂ + 10H₂O → 4Fe(OH)₃ + 8H⁺
(水酸化鉄、さらに脱水して酸化鉄Fe₂O₃へ)
酸化鉄は水に溶けず、海底に沈殿する。これが縞状鉄鉱層(BIF:Banded Iron Formation)だ。地球でも24億年前前後に大規模なBIFが形成され、酸素の証拠となっている。
ルブラリスでも同様のプロセスが起きた。約10億年間(35〜25億年前)、海洋のFe²⁺が完全に酸化され尽くすまで、酸素は大気に蓄積しなかった。
(2)還元ガスとの反応
大気中にはCH₄(メタン)やH₂S、H₂などの還元性ガスが残っていた。酸素はこれらと反応して消費される:
CH₄ + 2O₂ → CO₂ + 2H₂O
H₂S + 2O₂ → SO₂ + H₂O
これらの反応が進むまで、やはり酸素は蓄積できない。
やがてFe²⁺と還元ガスが枯渇すると、酸素は行き場を失い、大気へ溢れ出した。CO₂は徐々に減少し、O₂が増加した。しかし重要なのは、CO₂は決して地球のように0.04%まで下がらなかったという点だ。
なぜCO₂が高止まりしたのか?
理由は三つある:
(1)温室効果の必要性
前回で計算したように、平均気温30°Cを維持するには、CO₂が約4〜5%必要だ。これより下がれば、惑星は凍結する。光合成でCO₂を消費しすぎると、自らの生存環境を破壊することになる——これが下限を作った。
(2)火山活動による供給
潮汐加熱により、火山活動は地球の約10倍活発だ。年間2〜3 GtのCO₂が常に大気へ供給される。光合成による消費と、火山による供給が釣り合った点が、平衡濃度となる。
(3)風化作用の飽和
CO₂は岩石との風化反応で除去される:
CO₂ + CaSiO₃ → CaCO₃ + SiO₂
しかしこの反応速度は温度とCO₂分圧に依存する。ある程度高いCO₂濃度では、風化速度が頭打ちになる(反応速度論的飽和)。
これら三つの要因で、CO₂は4%前後で安定した。
酸素の蓄積は、生態系を激変させた。
(1)嫌気性生物の衰退
初期の化学合成生物(メタン生成菌など)の多くは、酸素に触れると死ぬ。酸素は彼らにとって猛毒だった。活性酸素種(・OH、O₂⁻など)がDNAやタンパク質を破壊するからだ。
生き残ったのは、深海や堆積物の中など、酸素の届かない場所へ逃げた種だけだ。彼らは現在でも、そうした「嫌気的ニッチ」で細々と生きている。
(2)好気呼吸の進化
一方、酸素を利用する新しい代謝系が進化した——好気呼吸だ:
C₆H₁₂O₆ + 6O₂ → 6CO₂ + 6H₂O + 大量のATP
グルコース1分子から、嫌気呼吸(発酵)では2分子のATPしか得られないが、好気呼吸では約36分子ものATPが得られる。18倍のエネルギー効率だ。
これは革命だった。好気呼吸を持つ生物は、同じ量の食物から遥かに多くのエネルギーを取り出せる。より活発に動き、より速く成長できる。彼らが生存競争を制した。
(3)真核生物の誕生
地球では約21億年前、真核生物が誕生した。真核生物とは、核膜で囲まれた核を持つ細胞の生物——私たち動植物を含む、複雑な生物すべてだ。
真核生物の起源は「細胞内共生説」で説明される。ある古細菌が、好気呼吸を行うバクテリアを飲み込んだ。しかし消化せず、体内に住まわせた。バクテリアは宿主にエネルギー(ATP)を供給し、宿主はバクテリアに保護と栄養を与える——相利共生だ。
やがてこのバクテリアは「ミトコンドリア」となり、細胞の「発電所」として機能するようになった。すべての真核生物は、このミトコンドリアを持っている——これが約20億年前の共生の証拠だ。
ルブラリスでも、同様のプロセスで真核生物が誕生したと考えられる。時期は地球より若干早い、22〜20億年前頃だろう。理由は光合成が早く進化し、酸素蓄積も早く始まったためだ。
こうして、ルブラリスの海は緑ではなく黒く染まった。メラノバクテリアが、海洋表層を覆い尽くした。近赤外の光を吸収し、OH⁻を電子源として、酸素を吐き出し続けた。
大気のCO₂は徐々に減少し、30%から4%へ——しかしそこで止まった。温室効果の壁だ。これ以上減らせば、惑星は凍りつく。同時に、酸素は0%から5%へ上昇した。地球の21%より体積比では地球(21%)より低いが、全圧4.5気圧ゆえ分圧では ppO₂=0.225 atm と地球海面(0.21 atm)並みで、好気呼吸には十分である。
むしろ、これ以上になると次の副作用が急に強まる。
燃焼・火災リスクの増大:分圧が上がるほど可燃物の発火・延焼は加速する。8〜10%O₂(ppO₂=0.36〜0.45 atm)域では、乾燥期や粉塵環境で小さな火花が大火になりやすい。ルブラリスは高CO₂(4%)と高湿潤で消炎・冷却されがちとはいえ、上限を押し上げれば釣り合いが崩れる。
生体の酸化ストレス:高ppO₂は反応性酸素種(ROS)の生成を押し上げ、色素・膜脂質・ヘム酵素の酸化損傷が増える。発光器官や視覚受容体のような高代謝組織はとくに脆い。
光化学の暴走:赤色矮星のフレアUVと高O₂が重なると、成層圏O₃は増える一方、対流圏では光化学スモッグ(過酸化物・NOx連鎖)が立ちやすくなる。通信・発光行動に霧散乱ノイズを増やす要因になる。
したがって、O₂=5%(ppO₂≈0.225 atm)は「好気代謝を十分満たしつつ、燃焼・酸化・光化学の暴走を抑える実用的な上限」になる。
3. 陸上進出の条件
メラノバクテリアは海洋を支配し、大気組成を変え、真核生物を生み出し、複雑な生態系を築いた。しかし、陸地は依然として不毛だった。岩と砂の荒野——それが陸の姿だった。生命が海から陸へ進出するには、まだ決定的な障壁が残っていた。
陸上進出の最大の障害は、紫外線(UV)だ。
主星セファエルはM型赤色矮星で、可視〜近赤外の全体光量は地球の約25%相当しか届かないが、静穏時でも短波長UVは少量ながら存在し、さらにフレアでFUV/NUV(遠紫外/近紫外)の比が跳ね上がる。フレアは小規模なものが日常的(数時間〜数日おき、TOAで平常の10~100倍)、稀に大規模(1000倍級)は月〜年スケールで起こる。
UVは生命にとって猛毒だ。DNAを直接破壊し、細胞を殺す。特にUV-C(200〜280 nm)とUV-B(280〜315 nm)は、核酸の二重らせんを切断する。海中では水自体の吸収と散乱で数十cm〜数m潜れば急減衰するが、陸上では遮蔽物がない。
オゾン層の形成(22〜20億年前)
救いは、酸素自身がもたらした。大気中の酸素分子(O₂)が、太陽紫外線を浴びると、酸素原子(O)に分解される:
O₂ + UV(< 242 nm) → 2O
生成した酸素原子が、別の酸素分子と結合すると、オゾン(O₃)ができる:
O + O₂ + M → O₃ + M
(Mは第三体:N₂など、反応の運動量を受け取る分子)
生成したO₃は200〜315 nmのUVを強く吸収して分解し、再びO₂とOに戻る。
O₃ + hν(200〜315 nm) → O₂ + O
生成と分解がつり合う層が成層圏(高度20〜40 km)にでき、それがオゾン層だ。UVCは事実上地表に到達しなくなり、UVBも大幅に減衰する。
ルブラリスでは、大気O₂が約5%(全圧4.5 atm → 分圧ppO₂≈0.225 atm)に達した頃(22億年前前後)、有効なオゾン層が成立した。M型星は静穏時のUVCが少ないためO₃生成光子が地球より弱めだが、フレア供給のUVが平均的な光化学収支を補う。その結果、地表の平常時UV-Cはほぼゼロ、UV-Bは地球の曇天〜薄曇り程度に抑えられる。
ただし、フレア時は別だ。強いフレアや荷電粒子イベントはNOx(窒素酸化物)を生成してO₃破壊の触媒になり、数時間〜数日のスパイクで地表UV-Bが平常の数倍に跳ね上がることがある。成層圏O₃の回復は光化学の時定数で数日〜数週間。この“危険日”を前提に、ルブラリスの生物は海表層で鍛えられた防御一式を、陸上でも流用した。要点は三つ。
この“危険日”を前提に、ルブラリスの生物は海表層で鍛えられた防御一式を、陸上でも流用した。要点は三つ。
(1)DNA修復酵素の高活性化
- 光回復酵素(photolyase):UV損傷を光エネルギーで直接修復
- ヌクレオチド除去修復(NER):損傷部位を切り出して再合成
- 地球生物の5〜10倍の酵素濃度
(2)UV吸収色素の蓄積
- カロテノイド(赤〜黄色):UV-Aを吸収
- フラボノイド相当:UV-Bを吸収
- 細胞表層に高濃度蓄積 → 内部のDNAを守る
(3)冗長性の確保
- DNA複製エラー率:地球の1/10(高精度ポリメラーゼ)
- 多重バックアップ:重要遺伝子を複数コピー保持
- 休眠機構:フレア検知(急激な光量変化)で即座に代謝停止
これらの防御機構は、海表層で進化したものだ。ルブラリスの海洋生物は、常にフレアに曝露されてきた。その結果、地球の陸上植物よりさらに強力なUV耐性を、最初から持っていた。
オゾン層が形成された時点で、ルブラリスの生物にとって陸上のUVは、「致命的」から「管理可能」な脅威へと変わった。
最初の陸上生命は、マングローブ様の群体植物だ。
潮の満ち引きがある海岸——満潮時には海水に浸かり、干潮時には空気に晒される場所。ここに、海藻から進化した生物が根を下ろし始めた。彼らは地球のマングローブと似た戦略を取った。
根:海水・底泥(Mn豊富)に浸かる
葉:気中で光合成
適応の詳細:
(1)気孔の獲得
気中での光合成には、気孔が不可欠だ。気孔とは、葉の表面にある微小な穴で、ガス交換を行う。
【気孔での交換】
内部 → 外部:O₂、H₂O(蒸散)
外部 → 内部:CO₂
当時の大気CO₂濃度は約10%だった(現在は4%)。地球の0.04%と比べれば250倍もある。しかしこれは両刃の剣だった。
高CO₂環境での気孔制御:
光合成時:気孔を開く → CO₂取り込み、O₂排出
呼吸時(夜間):気孔を大きく開く → 過剰CO₂の積極排出
フレア時:気孔を閉じる → UV遮断(クチクラ層で)
夜間の気孔開放が重要だった。昼間の光合成で内部CO₂は下がるが、夜間の呼吸で急速に上昇する。これを排出しないと、細胞質のpHが下がり(CO₂→H₂CO₃→H⁺)、酵素が失活する。
(2)クチクラ層の発達
陸上では乾燥が脅威だ。葉の表面にワックス質の膜(クチクラ層)を張り、水分損失を防ぐ。同時に、UV遮蔽の役割も果たす。
(3)維管束の原型
水と養分を根から葉へ運ぶ管——維管束の原型が形成された。当初は単純な細胞の列だったが、やがて木質化(リグニン蓄積)して強度を増した。
色と外観:
葉:黒褐色(葉黒体の近赤外吸収、OH⁻酸化型)
幹:暗褐色(タンニン、塩分耐性)
根:灰黒色(底泥の酸化鉄を纏う)
海岸線は、黒い森に縁取られた。満潮時、森の下部は水に沈む。干潮時、黒い幹と根が露出し、主星の暗赤色の光を浴びる。
しかしマングローブ相当植物には、致命的な制約があった。根が常に海水に接していなければならないことだ。
海洋では、OH⁻酸化が完璧に機能した。pH 9〜10の海水中には、OH⁻が豊富にある。しかし陸上では違う。気孔から取り込めるのは水蒸気(H₂O分子)だけで、OH⁻イオンは存在しない。
つまり、内陸へ進出するには、電子源をOH⁻からH₂Oに切り替える必要があった。
地球では、H₂O分解のMn₄CaO₅クラスター獲得に約8億年かかった(嫌気的光合成から酸素発生光合成まで、35億年前→27億年前)。
なぜそれほど時間がかかったのか?
理由は二つある:
(1)酸化電位の差
OH⁻酸化(pH 9): +0.70 V
H₂O分解(pH 7): +0.82 V
pH 5(酸性側, チラコイド腔に相当):E ≈ +0.94 V
海でのOH⁻優勢条件から、陸上の光化学装置(PSIIに相当)では内部を酸性側に保つため、必要電位が+0.2〜0.25 Vほど厳しくなる。差は小さく見えても、触媒設計と電子供与体の“段取り”(段階的なプロトン共役電子移動:PCET)には決定的だ。
(2)触媒構造の複雑さ
地球のシアノバクテリアが獲得したMn₄CaO₅クラスターは、極めて複雑だ:
4個のMnイオンが特定の立体配置
1個のCaイオンが構造安定化
5個の酸素が架橋
周囲のタンパク質が精密に配置
この構造を「偶然」獲得するには、膨大な試行錯誤が必要だった——それが8億年という時間だ。
しかしルブラリスには、決定的なアドバンテージがあった。既にMn触媒の基本設計を持っていたのだ。
海洋で進化したOH⁻酸化のMn₂〜Mn₄錯体——これが土台になった。必要なのは「ゼロからMn触媒を発明する」ことではなく、「既存のMn触媒を改良する」ことだった。
段階1:基質の拡張(25億年前)
最初のマングローブ相当植物は、根を海水に浸けながら、葉を空気中に伸ばしていた。葉の細胞内には、海水由来のOH⁻がわずかに残っていたが、やがて枯渇する。
ある変異株が、H₂Oを「予備的な基質」として使える能力を獲得した。Mn錯体の活性部位に、わずかな変異(アミノ酸1〜2個の置換)が起きただけだ。
効率は悪かった。OH⁻の1/10程度の反応速度しか出ない。しかしOH⁻が枯渇した時、H₂Oを使って光合成を続けられる——これは生死を分ける差だった。
段階2:触媒の最適化(24〜23億年前)
H₂Oを基質にする株が生存競争で有利になると、次の改良が雪だるま式に積み上がる。
- Mn数の増加:Mn₂ → Mn₃ → Mn₄(協同的な電子移動)
- Ca導入:構造安定化(Mg → Caへの置換)
- 水素結合ネットワーク:H₂Oを活性部位に誘導・配向
- プロトンチャネル:生成したH⁺を効率的に排出
- 葉黒体のプロトンポンプ強化で“腔は酸性/ストロマは塩基性”の勾配を深く維持(必要電位の上昇に対応)
これらの改良は、「完全な新規発明」ではなく、「既存構造の漸進的改良」だった。地球のように8億年ではなく、わずか数百万〜2000万年で完成した。
段階3:OH⁻酸化系の廃棄(23億年前〜)
やがてH₂O分解の効率がOH⁻酸化を上回ると、OH⁻系は不要になった。遺伝子の一部が失われ、完全にH₂O分解専用の光合成系が確立された。
決定的な構造:Mn₄CaO₅クラスター
最終的に得られた構造は、地球のMn₄CaO₅と機能的同等。これは収斂進化の典型だ。H₂O→O₂という課題には、多核Mn+Ca+酸素架橋+PCET配管という解が、化学的に“最短”だからだ。違いは到達までの道程の短さ——ルブラリスはMn触媒をすでに持っていたため、時間を圧縮できた。
こうして、陸上植物は完全な水分解能力を獲得した。もはや海水に依存する必要はない。内陸への道が開かれた。
H₂O分解を獲得した植物は、河口や沼沢地へ進出した。しかし本格的な内陸進出は、さらに別の障壁に阻まれた——土壌のMnだ。
内陸の土壌には、Mnは豊富にあった。全球凍結時の超酸性雨で濃縮されたMnO₂が、3000〜10000 ppmも含まれていた(地球の土壌は200〜3000 ppm)。
しかし問題は、MnO₂は水に溶けないことだった:
MnO₂(固体、褐黒色)
溶解度:< 0.01 mg/L(ほぼ不溶)
植物が吸収できるのは、Mn²⁺(マンガンイオン)だけだ。MnO₂をMn²⁺に還元する必要がある:
MnO₂ + 4H⁺ + 2e⁻ → Mn²⁺ + 2H₂O
この反応には、酸性環境(pH 4〜5)と還元剤が必要だ。しかし土壌はpH 7〜8の中性〜弱アルカリ性。還元剤もない。植物単独では、土壌のMnをほとんど利用できなかった。
転機は、約18億年前に訪れた。沼沢地の植物の根に、土壌菌が感染した。
当初は寄生だった。菌が植物の炭水化物を奪う、一方的な搾取。植物は弱り、成長が鈍った。
しかしある菌株が、変異を起こした。有機酸(クエン酸、シュウ酸)を大量生産する能力を獲得したのだ。
例えば、シュウ酸ではこのような酸化反応が起こる。
C₂O₄²⁻ → 2 CO₂ + 2 e⁻ (シュウ酸 → 二酸化炭素)
これが MnO₂ の還元反応
MnO₂ + 4 H⁺ + 2 e⁻ → Mn²⁺ + 2 H₂O
と結びつくと、
MnO₂ + C₂O₄²⁻ + 4 H⁺ → Mn²⁺ + 2 CO₂ + 2 H₂O
つまり、シュウ酸が酸化されると同時にMnO₂が還元される。
考えうるメカニズムとして、シュウ酸が還元剤として、MnO₂を化学的にMn²⁺へと還元。。一方、クエン酸は生成したMn²⁺をキレートし、溶解度を維持し、植物の吸収を手助けする。
根圏で起きる一連のプロセス
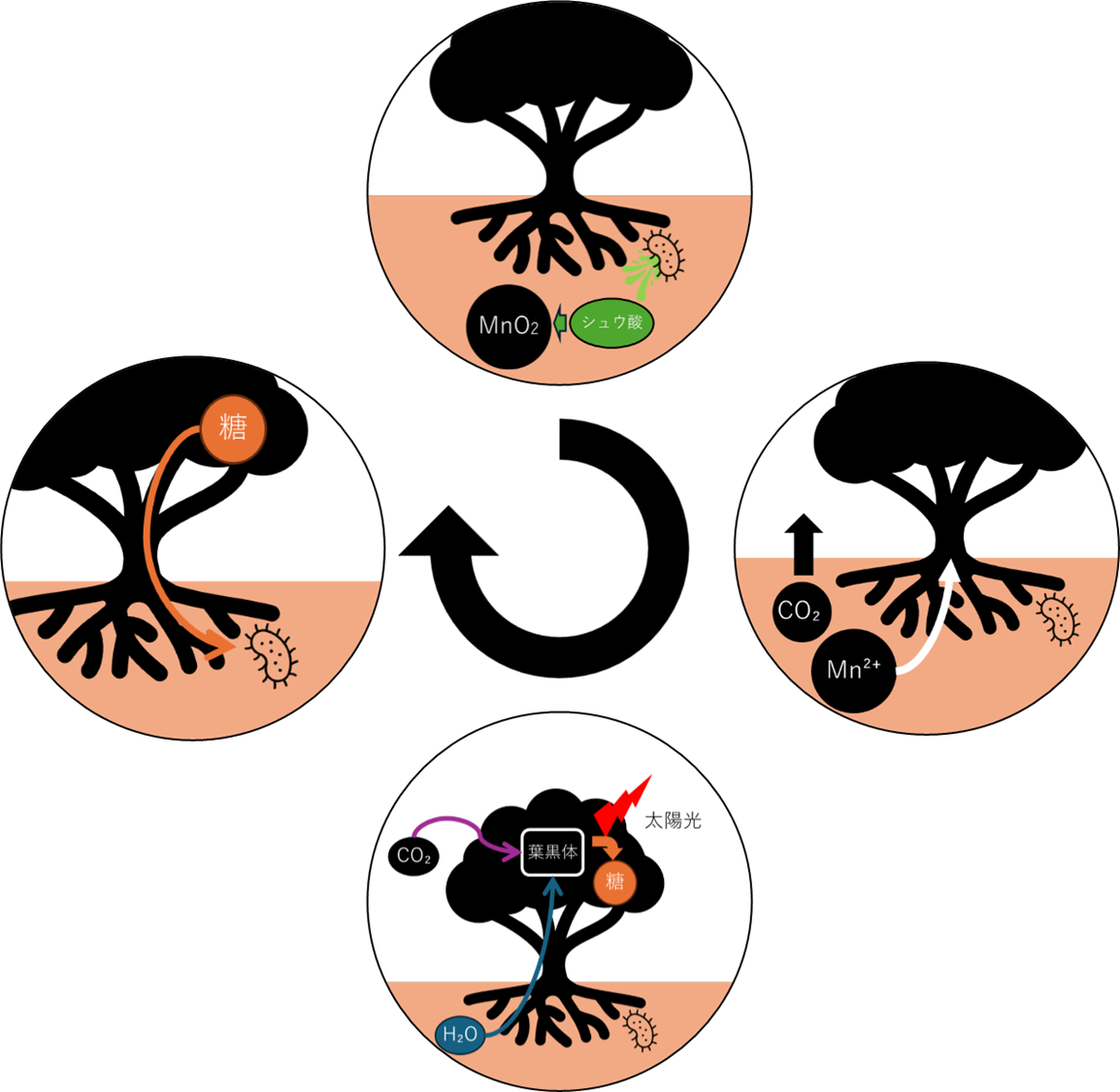
- 植物根のH⁺-ATPaseと共生菌の有機酸放出 → 根圏pH低下(4〜5)
- 有機酸と微生物代謝がMnO₂を還元 → Mn²⁺溶出
- 植物がMn²⁺吸収 → 光合成能力・抗酸化力が上がる
- 植物が共生菌へ糖供給を増やす → 菌がさらに有機酸を出す
これが「Mn動員菌根」の誕生——ルブラリスの陸上植物にとって、生存の絶対条件となる共生だった。
補記:なぜMnが主要養分なのか
地球では「肥料の三要素」——窒素(N)、リン(P)、カリウム(K)が作物生産を規定する。しかしルブラリスでは、Mnが第四の主要養分として加わる。
もちろん地球でもMnは必須だ。光合成の水分解触媒(Mn₄CaO₅クラスター)に使われ、葉乾物で20〜200 ppmほどあれば足りるのがふつうだ。
だがルブラリスでは桁が違う。
葉のMn濃度(代表値)
・地球の植物:20〜200 ppm
・ルブラリスの植物:500〜2000 ppm
(地球の代表的レンジ50〜150 ppm比で約10〜40倍)
理由は二つ、簡明だ。
(1)葉黒体密度が高い
主星光は地球比約25%。同じ一次生産を得るには、反応中心を物量で積むしかない。葉黒体(近赤外最適化)の実装密度は2〜3倍。各中心にMnが4原子必要であることを思い出せば、Mn需要も底上げされるのは自明だ。
(2)Mnが“周辺装置”でも主役
フレアと高UVは、常に活性酸素(ROS)を過剰に生む。これを掃くMn-SOD(マンガンスーパーオキシドディスムターゼ)や後段の過酸化物分解系の常時高発現が必須になる。内陸化に伴う木質化(リグニン化)や各種酸化還元酵素でもMnは要となる。
総合すると、ルブラリスの植物は500〜2000 ppmという“地球で言えば高Mn植物〜過蓄積域”に相当する濃度をふつうの運転点として要求する。これはもはや微量元素の域を超えた需要であり、栄養生態の現実としてN–P–K–Mnの四本柱が成立する。
(閑話休題)
内陸進出のごく初期、土壌はほぼ存在せず、分解者(菌類・細菌)も未発達で、養分循環はない。そんな中で唯一の“肥沃地”が、降灰の届く火山周辺だった。非晶質の火山ガラスやMn酸化物微粒子がパルス的(噴火年)に0.5〜5 t/ha、平年でも数kg/ha/年規模で供給され、根圏酸で可溶化しやすい新鮮Mnが継続的に補充される。Mn動員菌根を得たシダ相当植物は、この外部供給だけを梃子にまず火山帯で繁茂した。
火山近傍の帯状分布(長期平均)
- 半径 〜200 km:豊かな森林(樹高 30〜40 m 相当の群落潜在力/実際の優占は大型シダ 3〜5 m)
- 200〜500 km:疎林〜低木林(10〜20 m 潜在)
- 500 km超:草本優占〜荒地(1〜3 m)
やがて、落葉が積もりはじめる。
分解者がほとんどいないため、落葉・落枝(リター)は分解されず堆積する。数十万〜数百万年スケールで、火山周辺の台地や緩斜面には数 cm〜数十 cm の黒色マット層が形成され、C・N・P・Mn を含む有機物が“貯金”されていく。
大気酸素(ppO₂≈0.225 atm)が安定し、UVはオゾン層と表層色素で管理可能になった時期から、陸上性の菌類・細菌が徐々に多様化。リターに侵入し、セルロース・リグニン・タンニンへ作用する酵素群を獲得する。これにより腐植物質(フミン酸・フルボ酸)が生成し、初生土壌が成立する。ここでMn動員菌根は、根圏をpH 4〜5に酸性化してMnO₂→Mn²⁺の還元溶解を加速、可給性Mnを安定供給する役者へと定着する。
養分循環の立ち上がり
植物(近赤外優占の同化)
↓(落葉・落枝)
リター堆積(分解遅い → 次第に加速)
↓(菌類・細菌の分解)
腐葉土形成(フミン/フルボ)
↓(菌根の有機酸・キレート)
無機養分化(Mn²⁺、NH₄⁺、PO₄³⁻、K⁺)
↓(再吸収)
植物群落の維持・拡大
腐植層の成立により、群落のマンガン・窒素・リンの内部循環が回り始め、降灰や風化への依存は年々低下する。ただし、最初の繁栄は火山周辺が先行し、そこからリター→土壌→循環という順で内陸へ黒い群落が滲み出していった。
なお、初期の成長率は年 6〜10 cm 程度にとどまる。主星光は地球比25%と弱く、4.5 気圧で蒸散駆動が小さいため同化・水輸送が頭打ちになりやすい。さらに5 m超で導管の水頭限界と高密度大気の風動圧が効き、機械的安定性も制約となる。真正木部の発達(太い導管・二次成長)と気孔制御の高精度化が、次の飛躍に不可欠だった。ここから、火山帯発・黒いシダ林が、ついに“森”へ変わるための技術——樹木化——が始まる。
胞子植物の長い時代を終わらせたのは、胚と養分と堅牢な殻を一体化した「種子」という発明だった。発芽前から炭水化物と脂質を抱え、さらに高需要元素を前払いで蓄えることで、幼苗は貧栄養の礫地でも数週間は自力で立ち上がれる。殻は乾燥と紫外線と機械的衝撃を遮り、休眠は季節とフレアをやり過ごす時間的余裕を与える。しかも種皮の内外に菌根の胞子や菌糸を“同梱”するため、発芽と同時に根圏で有機酸が放たれ、Mn, N, Pを引き出す回路が即座に起動する。これらの利点が重なって、海岸や火山麓に限られていた黒い植生は、ついに内陸へと歩を進めた。
最初に大地へ本格的な森を築いたのは裸子植物相当である。彼らは厚い壁の仮導管と形成層を獲得し、年輪ごとに水柱の幹線を更新していく。深根は岩盤の割れ目まで潜り、外生型の菌根菌と組で鉱物を掘り起こす。その結果、樹高は三十〜五十メートルへと跳ね上がり、黒い針葉は主星の近赤外をほぼ吸い尽くす“光の終端層”を林冠に築いた。ただし、繁殖は風任せで、4.5 気圧の重い空気がもたらす厚い境界層と長い静穏夜が、花粉の到達を気まぐれにした。群落をまたぐ確実な遺伝子流動には、別の仕組みが要った。
その解として現れたのが被子植物相当、すなわち花と果実の戦略である。葉黒体が近赤外を貪欲に吸う黒い樹冠の中で、花弁は逆に近赤外を鏡のように反射する銀白——受粉者の複眼が最もよく見る帯域に、強烈なコントラストで「ここだ」と標識を出す。高濃度の蜜は不安定な光環境でも確実に訪花を誘い、粘着性の花粉は体表に絡みついて群落間を正確に橋渡しする。結実の段では、果皮が可視を吸って近赤外を透すことで、成熟した種子が散布者の視界に淡く“透ける”。消化を逃れる硬い種皮と、糞中でただちに発芽する菌根のコーティングが、長距離散布と即時定着を一つの連鎖に結ぶ。こうして受粉の精度と散布の距離が同時に引き上げられ、形態と生態の多様化は爆発した。




人気順のコメント(0)